|
Chevreulia and Cuatrecasasiella are diagnosed as herbs with opposite, distichous leaves, and a persistent pappus. Chevreulia contains six species primarily in austral South America with three reaching the northern Andean Cordillera and is diagnosed with fusiform achenes contracted into a filiform rostrum, barbellate pappus bristles, and biseriate, myxogenic trichomes approximately 40 µm long, with bulbous apical cells. Cuatrecasasiella is diagnosed as dioecious herbs with glabrous achenes; its two species represent closely related North/South disjuncts.
Historical Biogeography
A large portion of the South American Gnaphaliinae form a monophyletic group that displays a biogeographic pattern reflecting austral origins and subsequent south to north dispersal. The general pattern exhibited by the "Lucilia group" reflects the independent development of northern distributions from austral South American stock. Chionolaena is most diverse in the Atlantic Coastal mountains and adjacent habitats in eastern Brazil, with disjunct taxa in high-elevation sites in Colombia and Venezuela but with no representatives in the intervening Andean Cordillera. The distribution of the Achyrocline / Pseudognaphalium / Stenophalium clade is also congruent with an austral origin and subsequent invasion and radiation in the Andean Cordillera. Achyrocline has its greatest diversity in central and southern South America (Dillon & Sagástegui 1986a) but with secondary centers of speciation in Central America and Mexico (Nesom 1990a). Pseudognaphalium is most speciose in South America, followed by Mexico and the United States. The Gamochaeta clade also finds the greatest species diversity in austral South America with radiation both
in the Neotropics and elsewhere.
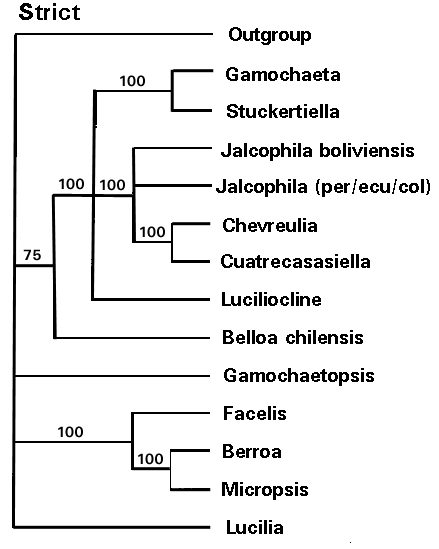
In all cladograms, the basal lineages in Lucilia are restricted either to Brazil or Argentina and sister group relations occur between taxa in the southern or northern Andean region. It can be hypothesized that as ancestral stocks were forced down slope by recurring glacial cycles, the fragmentation of pre-existing stocks through climate shift and uplift lead to northern and southern vicariance. The retreat of glacial conditions greatly changed the zonation in the alpine environments and the re-establishment of the Humboldt current formed an arid corridor to prevent distribution along the western versant. That Micropsis, Facelis and Berroa are highly derived members of the Lucilia stock is reflected by the results of Anderberg (1991) and the current analysis. These annual herbs represent diversification in seasonally dry habitats which developed repeatedly during Pleistocene and Holocene periods.
More recently, they have dispersed throughout Andean South America and ultimately North America, South Africa and Australia, often through the activities of commerce. In other genera, such as Luciliocline, the center of species diversity is in the Andean Cordillera, where typically occur species pairs with sister-species relationships between northern and southern Andean taxa (e.g. L. lopezmirandae and L. catamarcensis) suggesting vicariant speciation events.
The connection of South American taxa to those of New Zealand, Australia,
or Tasmania is a recurring pattern in many plant families. It is here
suggested that similar selection on independent stocks has lead to convergent
habits, but dissimilar floral morphologies. The relationship of South
American Gnaphalieae to those of Australasia will require further analysis
to support these hypotheses.
References (* added)
Anderberg, A. A. 1986. The genus Pegolettia (Compositae, Inuleae). Cladistics, 2: 158- 186.
________. 1988. The genus Anisothrix O. Hoffm. (Compositae-Inuleae). Bot. Jahrb. Syst. 109: 363-372.
________. 1989. Phylogeny and reclassification of the tribe Inuleae (Asteraceae). Canadian J. Bot. 67: 2277-2296.
________. 1991. Taxonomy and phylogeny of the tribe Gnaphalieae (Asteraceae). Opera Bot. 104: 1-195.
________. 1992. Cladistics of the Gnaphalieae, Additional data. Comp. Newsl. 20/21: 35.
________. 1994. Tribe Gnaphalieae. In K. Bremer, Asteraceae, Cladistics and Classification, pp. 304-364. Timber Press, Portland, Oregon.
________. & S. Freire. 1990. Jalcophila boliviensis, a new species
of South American Asteraceae (Gnaphalieae). Brittonia, 42: 138-141.
________.& ________. 1991. A cladistic and biogeographic analysis
of the Lucilia group (Asteraceae, Gnaphalieae). J. Linn. Soc. Bot.
106: 173-198.
Bentham, G. 1873. Notes on the classification, history, and geographical distribution of the Compositae. J. Linn. Soc. (Bot.) 13: 335-577.
Bremer, K. 1976a. The genus Relhania (Compositae). Opera Bot. 40: 1-85.
________. 1976b. The genus Rosenia (Compositae). Bot. Notiser, 129: 97-111.
________. 1978a. The genus Leysera (Compositae). Bot. Notiser, 131: 369-383.
________. 1978b. Oreoleysera and Antithrixia, new and old South African genera of the Compositae. Bot. Notiser, 131: 449-453.
________. 1987. Tribal interrelationships of the Asteraceae. Cladistics, 3: 210-253.
________. 1994. Asteraceae, Cladistics and Classification. Timber Press, Portland, Oregon. 752 pp.
Cabrera, A. L. 1958. El género Belloa Remy. Bol. Soc. Argent. Bot. 7: 79-85.
________. 1961. Observaciónes sobre las Inuleae-Gnaphalineae (Compositae) de América del sur. Bol. Soc. Argent. Bot. 9: 359-386.
________. 1978. Compositae. Flora de la Provincia de Jujuy. Colecc. Cient., Inst. Natl. Tecn. Agropec. 13(10): 275-288.
________. 1971. Compositae. Flora Patagónica. Colecc. Cient., Inst. Natl. Tecn. Agropec. 8(7): 1-451.
________. 1978. Compositae. Flora de la Provincia de Jujuy. Colecc. Cient., Inst. Natl. Tecn. Agropec. 13(10): 294-301.
Cronquist, A. 1950. A review of the genus Psilocarphus. Res. Stud. State Coll. Wash. 18(2): 71-89.
DeCandolle, A. P. 1838. Lucilia. Prodr. 7(1): 46.
Dillon, M. O. 1988. Generic limits and microcharacters in the South American Gnaphaliinae (Asteraceae-Inuleae).
Amer. J. Bot. Abst. 75: 456.
________. 1990. A tale of two genera: the implication of character choices and outgroup selection in cladistic analysis. Amer. J. Bot. Abst. 77: 129.
________. & A. Sagástegui A. 1986a. A new species of Achyrocline (Inuleae- Asteraceae). Phytologia, 59: 107-110.
________. & ________. 1986b. Jalcophila, a new genus of Andean Inuleae (Asteraceae). Brittonia, 38: 162-167.
________. & ________. 1990. Oligandra Less. revisited and the need for a new genus, Pseudoligandra
(Asteraceae: Inuleae). Taxon, 39: 125-128.
________. & ________. 1991a. Family Asteraceae: Part V. Tribe Inuleae. In, J. F. Macbride & Collaborators, Flora of Peru, Fieldiana: Botany, N.S. 26, 1-70.
________. & ________. 1991b. Synopsis de los géneros de Gnaphaliinae (Asteraceae-Inuleae) de Sudamérica. Arnaldoa, 1: 5-91.
Drury, D. G. 1970. A fresh approach to the classification of the genus Gnaphalium with particular reference to the species present in New Zealand (Inuleae-Compositae). New Zealand J. Bot. 8: 222-248.
________. 1971. The American spicate cudweeds adventive to New Zealand. New Zealand J. Bot. 9: 157-185.
Freire, S. E. 1986. Revisión del género Lucilia (Compositae, Inuleae). Darwiniana, 27: 431-490.
________. 1987. A cladistic analysis of Lucilia Cass. (Compositae, Inuleae). Cladistics, 3: 254-272.
________. 1989. Oligandra Less. is Lucilia Cass. (Compositae, Inuleae). Taxon, 38: 298-299.
________. 1993. A Revision of Chionolaena (Compositae, Gnaphalieae). Ann. Missouri Bot. Gard. 80: 397-438.
Hess, R. 1938. Vergleichend Untersuchungen ¸ber die Zwillingshaare der Compositen. Bot. Jahrb. Syst. 68: 435-496.
Hilliard, O. M., and B. L. Burtt. 1981. Some generic concepts in Compositae - Gnaphaliinae. J. Linn. Soc. Bot. 82:181-232.
Holub, J. 1976. Gamochaeta. In, Tutin, T. G., et al. (eds.), Flora Europaea, 4: 127.
Hooker, W. J. & G. A. W. Arnott. 1835. Contributions towards a Flora of South America and the Islands of the Pacific.
Comp. Bot. Mag. 1: 102-111.
Jansen, R. K., K. E. Holsinger, H. J. Michaels, & J. D. Palmer. 1990. Phylogenetic analysis of chloroplast DNA restriction site data at higher taxonomic levels: an example from the Asteraceae. Evolution, 44: 2089-2105.
________., H. J. Michaels, & J. D. Palmer. 1991. Phylogeny and character evolution in the Asteraceae based on chloroplast
DNA restriction site mapping. Syst. Bot. 16: 98-115.
Keeley, S. C., and R. K. Jansen. 1991. Evidence from chloroplast DNA for the recognition of a new tribe, the Tarchonantheae, and the tribal placement of Pluchea (Asteraceae). Syst. Bot. 16: 173-181.
Maddison, W. P., and D. R. Maddison. 1992. MacClade, Analysis of Phylogeny and Character Evolution, Ver. 3.0,
Sinauer Associates, Inc., Sunderland, Mass.
Merxmüller, H., P. Leins and H. Roessler. 1977. Inuleae--Systematic review. In V. H. Heywood et al. (eds.), The
Biology and Chemistry of the Compositae, pp. 577-602. Academic Press, London.
Morefield, J. D. 1992. Evolution and systematics of Stylocline (Asteraceae: Inuleae). unpubl. diss., 73 pps.
Nesom, G. L. 1989. New species, new sections, and a taxonomic overview of American Pluchea (Compositae: Inuleae). Phytologia, 67: 158-167.
________. 1990a. Taxonomy of Achyrocline (Asteraceae: Inuleae) in Mexico and Central America. Phytologia, 68: 181-185.
________. 1990b. Taxonomy of Gnaphaliothamnus (Asteraceae: Inuleae). Phytologia, 68: 366-381.
________.
1990c. An additional species of Gnaphaliothamnus (Asteraceae: Inuleae) and further evidence for the integrity of the genus. Phytologia, 69: 1-3.
________. 1994. Comments on Gnaphaliothamnus (Asteraceae: Inuleae). Phytologia, 76: 185-191.
*________. 2001. New combinations in Chionolaena (Asteraceae:
Gnaphalieae). Sida 19: 849-852.
Raven, P. H. and D. I. Axelrod. 1974. Angiosperm biogeography and past continental movements. Ann. Missouri Bot. Gard.
61: 539-673.
Remy, J. 1847. Compositae. In C. Gay, Historia Física y Política de Chile, Botánica, 3: 257-482.
Swofford, D. L. 1991. Phylogenetic Analysis Using Parsimony (PAUP), version 3.0s. Illinois Natural History Survey, Champaign.
Turner, B. L. 1977. Fossil history and geography. In V. H. Heywood et al. (eds.), The Biology and Chemistry of the Compositae, pp. 21-39. Academic Press,
London.
Appendix 1. Character and Data Matrix
A wide array of morphological characteristics have been examined, including habit, capitulescence type, and capitular and floral morphology (Dillon & Sagástegui 1991). Air-dried herbarium collections provided material for various types of microscopy. Material for light microscopy (LM) was treated with a wetting agent (Aerosol OT, 10%) and permanent slide mounts prepared using Hoyer's medium. Measurements were taken from the material prepared for LM. Material utilized for scanning electron microscopy (SEM) was mounted on stubs and gold coated without pretreatment. Voucher specimens are all housed at F unless otherwise noted.
1. Duration: (0) perennials, (1) annuals. 2. Lignification: (0) subshrubs, (1) herbs. 3. Leaf arrangement: (0) alternate, (1) opposite.
4. Leaf distribution: (0) evenly cauline, (1) basal rosette. 5. Leaf ranking: (0) non-distichous, (1) distichous. 6. Leaf
margins: (0) revolute, (1) plane, (2) involute. 7. Dorsal leaf surfaces: (0) non canaliculate, (1) canaliculate. 8. Leaf pubescence: (0) both tomentose, (1) abaxial tomentose, (2) adaxial tomentose. 9. Apical tuft on leaf: (0) absent, (1) present. 10. Foliar trichome apical cells: (0) swollen, (1) straight. 11. Capitulescences: (0) solitary, (1) glomerules, (2) spiciform, (3) panicles, (4) racemes. 12. Pedicles: (0) static, (1) elongating. 13. Capitular sexuality: (0) perfect (heterogamous), (1) polygamo-dioecious (heterogamous), (2) dioecious (homogamous), (3) perfect (homogamous). 14. Gynoecia (hermaphrodite): (0) fertile, (1) sterile. 15. Receptacles: (0) epaleate, (1) paleate. 16. Outer phyllaries and florets: (0) not enclosing, (1) enclosing. 17. Hermaphrodite florets: (0) 5-merous, (1) 4-merous. 18. Hermaphrodite style apices: (0) attenuate, (1) obtuse to rounded, (2) truncate. 19. Hermaphrodite styles dorsally: (0) pubescent or papillose, (1) glabrous. 20. Anther apical appendages: (0) isomorphic, (1) dimorphic. 21. Pappus bristles (pistillate): (0) acute to attenuate, (1) obtuse to rounded, (2) clavate. 22. Pappus bristles (hermaphrodite): (0) acute to attenuate, (1) obtuse to rounded, (2) clavate. 23. Trichome sizes: (0) elongate, (1) elongate-globose, (2) capitate-globose, (3) sessile. 24. Pappus condition: (0) deciduous, (1) persistent. 25. Achene shape: (0) oblong to oval, (1) rosulate. 26. Achene surface: (0) trichomes, (1) glabrous 27. Achenial trichome apices: (0) elongate, (1) capitate, (2) sessile. 28. Achenial trichome apical cells: (0) non-myxogenic, (1) myxogenic. 29. Achenial trichome apical cells: (0) untwisted, (1) twisted. 30. Achenial trichomes: (0) non-dehiscent, (1) dehiscent by cell splitting, (2) dehiscent by apical pores. 31. Achenial trichome bases: (0) enlarged myxogenic c-cell, (1) enlarged cells absent. 32. Achenial trichome types: (0) monomorphic, (1) dimorphic. 33. Trichome apical cells: (0) fused unequal, (1) fused equal, (2) unfused equal. 34. Carpopodium: (0) multiseriate, (1) 1-3-seriate. 35. Chromosome number: (0) x = 7, (1) x = 6.
Data Matrix Click here to access the data matrix image.
|








{kind=link}
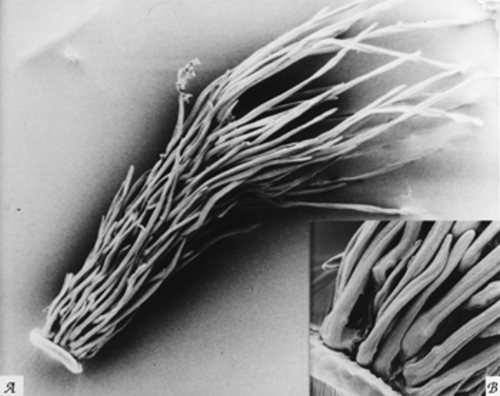
![Fig. 7. Belloa chilensis. A. Achene [1150 µm long]. B. Achene surface enlargement. C. Carpopodium enlargement. (Voucher: Teillier et al. 2020, F).](http://www.sacha.org/Gnaphalieae/images/Belloa_chilensis_web.jpg){kind=link}
![Fig. 8. Luciliocline longifolia. A. Achene [1280 µm long]. B. Achenial trichomes enlargement. (Voucher: Sagástegui et al. 12841, F).](http://www.sacha.org/Gnaphalieae/images/Lucilio_longifolia_web.jpg){kind=link}