
|
Jalca Formations of Northern Peru
Michael O. Dillon |
|
Andean Cordillera
The Andes comprise one of the most prominent structural features of South America. From initial orogeny dating to pre-Cretaceous and final uplift during the Plio-Pliestocene (no less than a million years ago), these mountains have been a dynamic pallet for Nature's processes for an extended period. They form a continuous, high elevation chain, largely unbroken for over 7500 kms along the Pacific side of South America, generally above 3000 m, between Venezuela and Tierra del Fuego. The interaction of elevation with climate and hydrology produce an exceedingly varied environmental mosaic ranging from arid intermontane valleys to very wet montane forests and varied alpine environments. The development of these features has undoubtedly influenced the differentiation of many elements of the South American biota. |
|
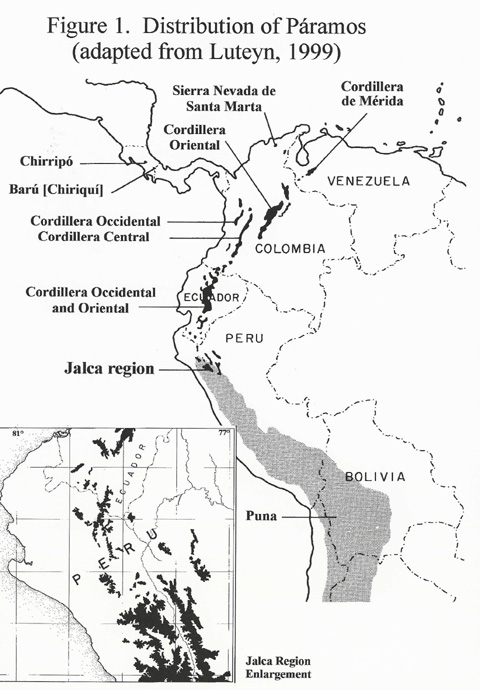
The interaction between high elevations and latitudinal range yield several types of specialized alpine environments broadly referred to as páramos (Cuatrecasas, 1968; Luteyn, 1999, Fig. 1). In northern Peru, between the páramos to the north and puna to the south, there exist transitional alpine communities above 3000 m known locally as jalca formations (Weberbauer, 1936, 1945) These formations are restricted to an area roughly bounded by 4.5-8° S latitude and 77-80° W latitude and are estimated to contain 420,000 hectares (Sánchez Vega, pers. comm.) or ca. 4200 km sq. While exacting definitions are lacking, jalca is usually interpreted as drier than páramo and wetter than puna (Bazán-Zurita et al., 1998; Luteyn, 1999; Sánchez, 1976). Luteyn (1999) has suggested that, generally, the jalca formations are more similar to the páramos of southern Ecuador. It remains an open question if this region is distinguished by either biotic or climatic parameters or if the region can be thought of as a more diverse and complex northern extension of the “altiplano” (Cabrera & Wallink, 1973). |
|
|
|
Huancabamba Deflection
While the main chain of the Andes is largely un-interrupted, in northern Peru (4°-6° S lat.) where there exists a pronounced discontinuity as the mountains bend from NW to NE and become fragmented, resulting in one of the lowest bisecting passes anywhere from Colombia to Terria del Fuego. This region is referred to as the Huancabamba Deflection (also termed Huancabamba Depression, Piura Divide, North Peruvian Low and in Peru "Depression de Huarmaca" for the district), so-named because of the change in orientation of the Andean chains (Fig. 1). Here, the Amazonian and Pacific slopes are separated by a pass, the Abra de Porculla, only slightly over 2000 m in elevation. |
|
The Huancabamba Deflection is only about 100 km wide and passes directly through the Departments of Piura, Cajamarca, to Amazonas in northern Peru. Cajamarca is the source of no fewer than a dozen cuencas (source basins) with rivers feeding both the Atlantic and Pacific versants. This region is geographically complex and marks where the primary Cordillera breaks up into a series of chains of small isolated ranges of low, dissected mountains with deep, dry valleys generally 1000-2000 m in elevation. It is delimited on its western edge by the Cordillera Huancabamba and the south flowing Río Huancabamba (SSE at from ca. 5°15" S, 79°27'W). The Rio Chotano flows north and meets the Río Huancabamba to form the southern arc of the eastward flowing Río Chamaya (ca. 6° S) which inturn joins the north-flowing Río Marañón and south flowing Río Chinchipe from southern Ecuador. |
|
The interior of the Upper Río Marañón and its tributaries in this area are dry, rain-shadow valleys that produce a sharp environmental contrast with the humid Amazonian forests farther east. Throughout the region, however, on both Amazonian and Pacific slopes, wet cloud forest forms at appropriate elevations, generally between 1500-2500 m, where ever the slopes trap sufficient moisture; above the forested zones, more open jalca vegetation formations cover the higher elevations. At lower elevations on the Pacific slopes, the montane vegetation gives way to thorn scrub and extremely arid deserts. Habitat heterogeneity and fragmentation in this region appears to have affected speciation, endemism, and overall biodiversity in both plants and animals. |
|
|
| |
|
Thus, the Huancabamba Deflection and the jalca formations, that occupy the upper-elevations within the region, form an extremely complex environmental mosaic, perhaps more varied than any other region of the Andes (Young & Reynel, 1997). The physiographic complexity of the Deflection region has led to generally poor understanding of the importance of the region both as a potential barrier to montane animal and plant lineages, and as a potential corridor between Amazonian and Pacific slope floras and faunas. Adding to the difficulty of interpreting distribution patterns in the Huancabamba Deflection are uncertainties regarding whether particular patterns are a reflection primarily of the present environmental mosaic; or if present-day distributions reflect deeper historical events that might be discovered by appropriate phylogenetic and historical biogeographic approaches. Both scales of inquiry are relevant to gaining a comprehensive insights into the processes that have shaped these communities. The jalca formations above the village of Bolivar in extreme northeastern La Libertad, represent some of the most isolated pockets of alpine vegetation on the eastern escarpment.
|















