 |
Griseliniaceae |

| |
Griseliniaceae |
A version of the following information has been submitted to be published in an upcoming volume of The Families and Genera of Vascular Plants, Dillon, M.O. xxxx. Griseliniaceae. Pages xx-xx. In: K. Kubitzki (ed.), The Families and Genera of Vascular Plants. Vol. Xx. Springer Verlag, Berlin. Do not cite!
Griseliniaceae J.R. Forst. & G. Forst. ex A. Cunn.
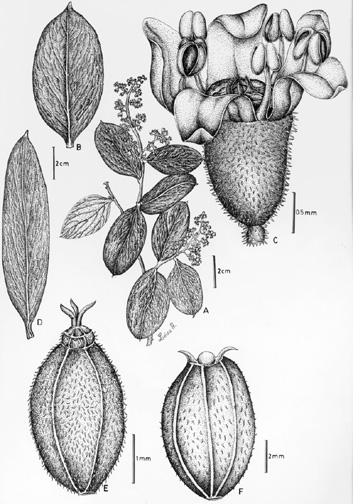
Evergreen, dioecious trees, shrubs, or lianas. Stems erect to scandent or climbing, or occasionally epiphytic, glabrous to hispidulose, rarely winged. Leaves simple, coriaceous, entire to dentate, dentate-spinose or dentate, alternate to subspiral or subopposite; subsessile or petiolate. Petioles rugulose, glabrous or hispidulose, exstipulate, the bases dilated and subsheathing. Blades variable, broadly-ovate to lanceolate, elliptic, oblong, or rhomboid, conspicuously asymmetrical or occasionally symmetrical, coriaceous, basally oblique to equal, rounded, obtuse, truncate, or cuneate, apically rounded to retuse, or acute, attenuate, or acuminate, the abaxial surface penninerved or prominently 3-5-nerved from near the base, reticulate, minutely pellucid-puncticulate, the adaxial surface smooth (drying rugulose), nitid, the margins revolute or plane, strictly entire to dentate or spinose or dentate-spinulose, occasionally subentire with only the apex minutely 2-3-fid, the teeth spinose to mucronulate. Inflorescences racemose to paniculate, terminal or axillary, subglabrous to hispidulose or glandular, drying nigrescent. Bracteoles caducous. Flowers unisexual, actinomorphic, 5-merous; hypanthium present. Sepals squamellate, margins ciliolate to glabrous, caducous. Petals 5, free, occasionally absent, subimbricate, oblong to ovate, reflexed at anthesis, caducous, margins ciliolate to glabrous. Stamens 5, antisepalous, the filaments erect, attached outside of lobulate epigynous disc. Anthers bilocular, dorsifixed, dehiscence longitudinal. Gynoecium 3-carpellate, syncarpous, ovary 2-locular (only one locule ovuliferous), inferior; styles 3, simple, terminal, free to connate basally, subulate, divergent or recurved; placentation apical; ovules pendulous; anatropous. Fruit fleshy, baccate or coriaceous, drupaceous, oval to oblong, glabrous to hispidulose-purberulent; 1-seeded. Seeds with copious endosperm; embryo minute; x = 9 (n = 18).
Seven spp., trans-Pacific disjunct (Argentina, Brazil, Chile, New Zealand).
VEGETATIVE MORPHOLOGY AND ANATOMY. All members of the family are woody, ranging from small trees or large shrubs reaching a maximum height of 10-20 m to shrubs seldom over 2 m. The growth habit is typically erect and self-supporting, but epiphytic and climbing habits are recorded. In those, the juvenile individual begins as an epiphyte and sends roots down to the ground to establish itself as a separate tree. Stomata are generally confined to the abaxial surface and cyclocytic. An adaxial hypodermis is present, the mesophyll contains sclerencymatous idioblasts, and minor leaf veins are without phloem transfer cells. Secretory cavities are absent, a cork cambium is present, nodes are tri-lacunar or multi-lacunar. Secondary thickenings develop from a conventional cambial ring, the xylem contains vessels with horizontal, scalariform end-walls (fibers with bordered pits), and the wood parenchyma is apotracheal (Li & Chao 1954).
INFLORESCENCES. The individual flowers are arranged in open, racemose to paniculate inflorescences which are borne either terminal or axillary on branchlets. The peduncles are typically subglabrous to hispidulose or glandular, nigrescent; bracteoles caducous.
FLORAL MORPHOLOGY. Flowers are sexually dimorphic. The staminate flowers have a campanulate hypanthium and are always petaliferous. The petals are yellow to greenish-yellow, reflexed at anthesis, and quickly caducous. Anthers are opposite the sepals and alternating with the petals. The pistillate flowers in a few species do not develop petals. The epigynous disc is not unlike those found in Araliaceae.
EMBRYOLOGY. The ovule morphology has only been investigated in one species and found to be unitegmentary and crassinucellate (Philipson 1977).
POLLEN MORPHOLOGY. Pollen shed as single grains. Pollen grains 3 - aperturate; colporate, ellipsoidal with a complete tectum with striate surfaces (Ferguson & Hideux 1978; Tseng 1980). The first appearance in the fossil record is from New Zealand in Miocene (Mildenhall 1980).
KARYOLOGY. Only one reported chromosome count of n = 18 exists for the family (Federov 1969)
POLLINATION. (*no data available)
REPRODUCTIVE SYSTEMS. All members of the family are dioecious. The gynoecium of staminate flowers is absent and no rudimentary stamens are found in the pistillate flowers. Schlessman et al. (1990) suggested that the dioecious condition in the Apiales arose from ancestral andromonoecy, but this has not been confirmed for this family.
SEED. (*no data available)
DISPERSAL. The presence of fleshy fruits may suggest some type of animal mediated dispersal, however, direct observations are lacking.
PHYTOCHEMISTRY. Jensen and Nielsen (1980a, b) investigated five Griselinia species and isolated the iridoid glucoside, griselinoside, from G. littoralis, G. lucida, G. ruscifolia, and confirms that iridoids are they are elaborated in both New Zealand and South American species. Reports indicate that the family is not cyanogenic and lacks proanthocyanidins and ellagic acid.
SYSTEMATICS. The family is monogeneric with the seven species of Griselinia. The history of familial placement has been complex. Early accounts suggested placement in families such as Euphorbiaceae and Juglandaceae (see Taubert 1892). Endlicher (1850) placed Griselinia in the Araliaceae and J. D. Hooker (1867) was the first to position it as a member of the Cornaceae. Taubert (1892) accepted the placement in the Cornaceae and established two sections, Decostea Taub. and Eugriselinia Taub. Takhtajan (1980) treated Griselinia as a monogeneric family and allied it with Garrya (Garryaceae). Thorne (1968), Dahlgren (1980), and Cronquist (1981) considered it an anomalous member of the Cornaceae. Thorne (1992, 2001) has placed it as a monogeneric family with relationships outside of the Cornales. Dillon & Muñoz-Schick (1993) monographed Griselinia, considering it a monogeneric family and described a new northern Chilean desert species.
AFFINITIES. Philipson (1967) was the first worker to question the placement of Griselinia in the Cornaceae, contending that it shared many features with the Araliaceae and Escalloniaceae, including geographic distribution, vegetative morphology, nodal anatomy, and the floral vascular system. Philipson (1977) suggested Griselinia be placed in the informal group Unitegminae, a complex of orders and families (including Cornales, Araliales, Escalloniaceae, and Pittosporaceae) possessing ovules with a single integument, a character shared with Aralidium. (Aralidiaceae, Philipson & Stone 1980). Eyde (1988) also recognized that Griselinia was misplaced in the Cornaceae and suggested familial rank.
Jensen and Nielsen (1980a) found the co-occurrence of the iridoid glucoside, griselinoside, in Aralidium, Toricellia and Griselinia, but the same compound was also isolated from Verbena (Damtoft et al. 1983). This co-occurrence was attributed to a common, homologous biosynthetic apparatus in these species (Jensen & Nielsen 1980b).
Xiang et al. (1993) and Xiang and Soltis (1998) examined rbcL sequence data and suggested Griselinia be removed from the Cornaceae and grouped it with Aralidium and Toricellia and more distantly with Aralia, Pittosporum, and Sanicula. Plunkett et al. (1996) also examined rbcL sequences and confirmed that the Apiales form a monophyletic group with Pittosporaceae along with Griselinia, Melanophylla (Malagasy endemic), Toricellia (E. Himalayas to W. China) and Aralidium (W. Malaysia).
DISTRIBUTION AND HABITATS. The family has a trans-Pacific disjunct distribution between austral South America and New Zealand. All the South American species are confined to Chile and adjacent Argentina with an additional disjunction to southeastern Brazil. The local environments vary greatly, from mesic, evergreen rain forest environments of austral Chile (sea level-1300 m) to dry coastal quebradas of the Atacama Desert in northern Chile (400 m). Southern Brazilian populations are found in Araucaria forests and range north to remnants of the Atlantic coastal forest (400-2700 m). The New Zealand species are distributed from near ocean sites to montane forests of North and South Islands.
Genera of the Griseliniaceae
1. Griselinia J. R. Forst. & G. Forst.
Griselinia J. R. Forst. & G. Forst., 1775, non Scopoli, 1777.
Decostea Ruiz & Pav., 1794.
Pukateria Raoul, 1844.
Selected Bibliography
Cronquist, A. 1981. An integrated system of classification of flowering plants. Columbia University Press, New York.
Dahlgren, R.M.T. 1980. A revised system of classification of the angiosperms. Botanical Journal of the Linnean Society 80: 91-124.
Damtoft, S., S. R. Jensen, & B.J. Nielsen. 1983. Biosynthesis of the iridoid glucosides cornin, hastatoside, and griselinoside in Verbena species. J. Chem. Soc. Perkin Trans. 1: 1943-1948.
Dillon, M.O., Muñoz-Schick, M. 1993. A revision of the dioecious genus Griselinia (Griseliniaceae) including a new species from the coastal Atacama Desert of northern Chile. Brittonia 45: 261-274.
Endlicher, S. L. 1850. Genera plantarum. Suppl. 5. (Griselinia, p. 16, n. 4562).
Eyde, R. H. 1987. The case for keeping Cornus in the broad Linnaean sense. Syst. Bot. 12: 505-518.
Federov, A. 1969. Chromosome numbers of flowering plants. Scientific Press,
Leningrad.
Ferguson, I. K., & M.J. Hideux. 1978. Some aspects of the pollen morphology and its taxonomic significance in Cornaceae sens. lat. Proc. IV Int. Palynol. Conf., Lucknow (1976-77) 1: 240-249.
Hooker, J. D. 1867. Cornaceae. Pp. 951. In: G. Bentham and J. D. Hooker (eds). Genera Plantarum 1(3).
Jensen, S. R. & B. J. Nielsen. 1980a. Iridoid glucosides in Griselinia, Aralidium and Toricellia. Phytochemistry 19: 2685-2688.
Jensen, S. R., & B.J. Nielsen. 1980b. The systematic position of Aralidium Miq. - A multidisciplinary study. 4. Iridoid glucosides. Taxon 29: 409-411.
Li, H., Chao, C. 1954. Comparative anatomy of the woods of the Cornaceae and allies. Quart. J. Taiwan Mus. 7: 119-136.
Mildenhall, D.C., 1980. New Zealand Late Cretaceous and Cenozoic plant biogeography: a contribution. Palaeogeogr., Palaeoclimatol., Palaeoecol. 31: 197-233.
Philipson, W.R. 1967. Griselinia Forst. fil. - Anomaly or link. New Zealand J. Bot. 5: 134-165.
Philipson, W.R. 1977. Ovular morphology and the classification of the dicotyledons. Plant. Syst. Evol., Suppl. 1: 123-140.
Philipson, W.R., Stone, B.C. 1980. The systematic position of Aralidium Miq. - A multidisciplinary study. 1. Introduction and floral and general anatomy. Taxon 29: 391-403.
Plunkett, G.M., Soltis, D.E., Soltis, P.S. 1996. Higher level relationships involving Apiales (Apiaceae and Araliaceae) based on phylogenetic analysis of rbcL sequences. Amer. J. Bot. 83: 499-515.
Schlessman, M.A., Lloyd, D.G., Lowry, P.P. 1990. Evolution of sexual systems in New Caledonia Araliaceae. Memoires of the New York Botanical Garden 55: 105-117.
Takhtajan, A. 1980. Outline of the classification of flowering plants (Magnoliophyta). Bot. Rev. (Lancaster) 46: 225-359.
Taubert, P. 1892. Revision der Gattung Griselinia. Bot. Jahrb. Syst. 16: 386-392.
Thorne, R. F. 1968. Synopsis of a putatively phylogenetic classification of the flowering plants. Aliso 6(4): 57-66.
Thorne, R. F. 1992. Classification and Geography of the Flowering Plants. Bot. Rev. (Lancaster) 58: 225-348.
Thorne, R.F. 2000 (2001). The classification and geography of the flowering plants: dicotyledons of the class Angiospermae (subclasses Magnoliidae, Ranunculidae, Caryophyllidae, Dilleniidae, Rosidae, Asteridae, and Lamiidae). Bot. Rev. 66: 444-647.
Tseng, C. C. 1980. The systematic position of Aralidium Miq. - A multidisciplinary study. 3. Pollen Morphology. Taxon 29: 407-409.
Xiang, Q.Y., D.E. Soltis, D.R. Morgan, & P.S. Soltis. 1993. Phylogenetic relationships of Cornus L. sensu lato and putative relatives inferred from rbcL sequence data. Annals of the Missouri Botanical Garden 80: 723-734.
Xiang, Q.Y. & D.E. Soltis. 1998. RbcL sequence data define a cornaceous clade and clarify relationships of Cornaceae sensu lato. Pp. 123-137. In: D.E. Boufford & H. Ohba (eds.). Sino-Japanese flora, its characteristics and diversification. University Museum, University of Tokyo, Bulletin 37. Tokyo.
Dillon, M.O., Mu±oz-Schick, M. 1993. A revision of the dioecious genus Griselinia (Griseliniaceae) including a new species from the coastal Atacama Desert of northern Chile. Brittonia 45: 261-274.
| [ABSTRACT: The genus Griselinia (Griseliniaceae) is revised and a new species, Griselinia carlomunozii, is described from the coastal quebradas in the Atacama Desert of northern Chile. An illustration, a discussion of its possible relationships, a key, and descriptions of all species are provided.] |
| Griselinia carlomunozii is based upon: TYPE: CHILE. Antofagasta (Regi¾n II): Prov. Antofagasta. Quebrada Rinconada,
. 5 km N of Caleta Paposo, (24░56'W, 70░30'W), 240-440 m, 4 Nov 1988, [staminate individual], M. O. Dillon & D. Dillon 5864 (Holotype: F; isotypes: GH, MO, NY, SGO).
|
 M. Dillon with G. carlomunozii at type locality, 4 Nov 1988 |
 Close-up of staminate flowers |

[Retorno al Homepage]
{kind=link}